Recent News
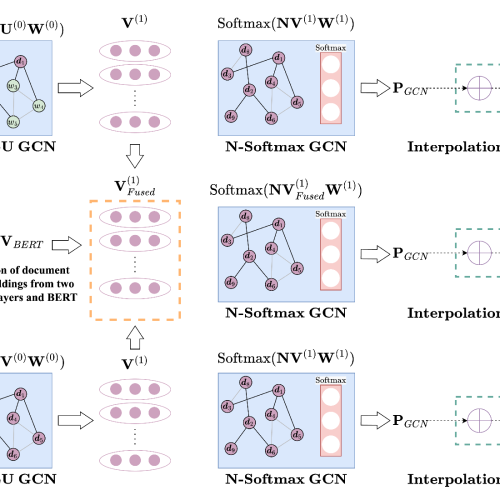
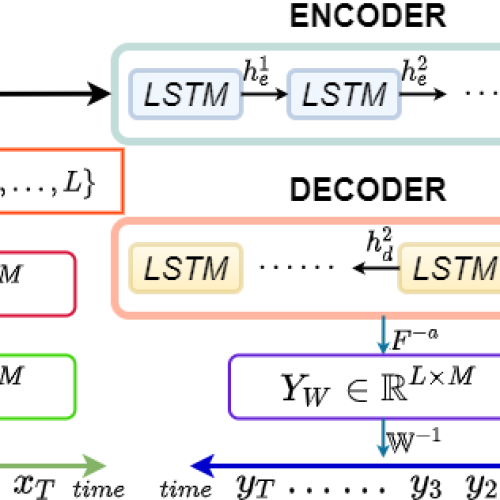




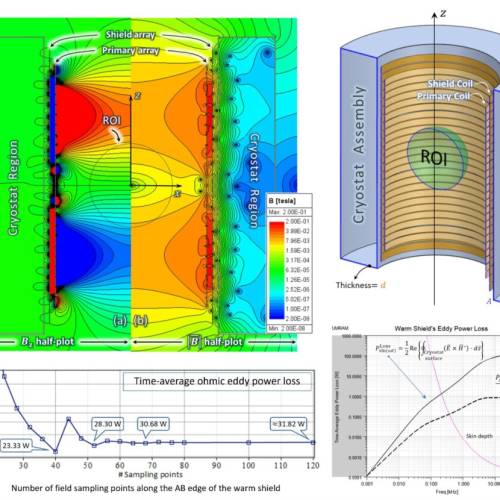

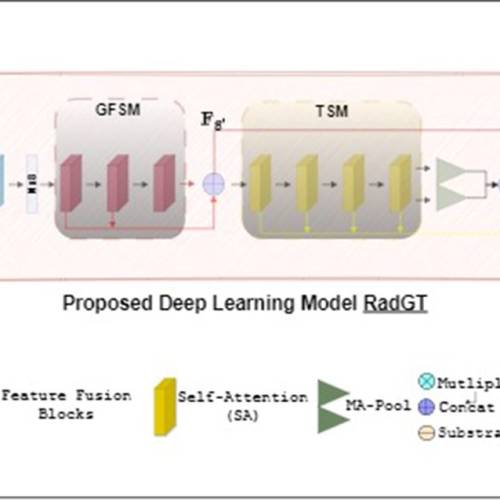
Latest Publications


80+
Grad Students
100+
Alumni
15
Faculty Members
10
Post Doc & Staff
600+
Publications
40+
Awards
50+
Projects
100+
Conferences & Seminars